CARIBBEAN
The Caribbean region is a Conservation International hotspot of biological diversity (http://www.biodiversityhotspots.org/xp/hotspots/caribbean/Pages/default.aspx), yet detailed processes that generate species diversity on the islands are complex and poorly understood. Our NSF funded project (DEB-1050187-1050253) aims to test hypotheses about how the age and dispersal ability of lineages and the geological history of islands interplay to generate biodiversity hotspots. To do so we will use multiple lineages of arachnids - spiders and relatives, including extant species and fossils.
The Caribbean region is a Conservation International hotspot of biological diversity (http://www.biodiversityhotspots.org/xp/hotspots/caribbean/Pages/default.aspx), yet detailed processes that generate species diversity on the islands are complex and poorly understood. Our NSF funded project (DEB-1050187-1050253) aims to test hypotheses about how the age and dispersal ability of lineages and the geological history of islands interplay to generate biodiversity hotspots. To do so we will use multiple lineages of arachnids - spiders and relatives, including extant species and fossils.

The Caribbean archipelago consists of some 7000 islands and islets, stretching over 4000 km.
Geological history of the Caribbean plays a central role in the formation and structure of biodiversity. Key parameters include geological mechanism of island origin (e.g. - Darwinian or fragment), proximity to major landmasses, island size and island age. The Caribbean islands provide a context in which to compare differences in these geological variables; the region includes over 7000 islands and islets, forming an island chain over 4000 km long, supporting exceptionally diverse ecosystems ranging from cactus deserts to rainforests. The larger islands also have impressive elevational range with rich habitat variety. There are three broad categories of islands in the Caribbean (Fig. 1): (1) ‘Darwinian’ islands (Gillespie and Roderick 2002) that have formed de novo and have always been surrounded by sea; (2) Fragment islands produced via fragmentation of continental crust and reached their current position by tectonic plate movement; and (3) Platform islands, low elevation islands that have regularly been submerged with changing sea-level. Also, various components of the Greater Antilles have fragmented and rejoined, e.g. NW Hispaniola is a geological relative of Cuba, while SW Hispaniola is not (Iturralde-Vinent & MacPhee 1999).
Species diversification patterns are expected to differ between the above island types. For example, Darwinian islands provide a clean slate to colonists where species richness increases initially through immigration and later through formation of neo-endemics. For highly isolated islands, adaptive radiation may fill empty niches (Gillespie & Roderick, 2002, Gillespie 2004). Fragment islands ought to be species rich upon birth, with ecological space initially filled. Species richness then changes after fragmentation as a function of extinction, colonization, and in situ speciation (Gillespie & Roderick, 2002). The cyclically submerging and reemerging of platform islands as sea levels change causes extinctions and thus limits diversity. The different geological histories of islands lead to different predicted patterns of diversity and types of dispersers they harbor. Given equal age, size, and isolation, we predict that Darwinian islands will hold the highest relative diversity of endemic species of intermediate to good dispersers, but will lack lineages that are very poor dispersers. Fragment islands will have the highest diversity, and be characterized by lineages of poor dispersers that were present prior to fragmentation. Platform islands, with their biota repeatedly wiped out, will be relatively species poor, have low endemism and inhabited by relatively good dispersers. Adding variation due to island isolation, age and size, we expect a highly mosaic landscape of species diversity and distributions.
The Caribbean as an opportunity to test island–continent dispersal dynamics. The paradigm of island biogeography has long seen biotic interchange as unidirectional with larger landmasses acting as sources and islands as sinks (MacArthur & Wilson, 1967; Wilson, 1961). No doubt this is a prominent pattern, however, Heaney (2007) and others (Bellemain & Ricklefs, 2008) foresee a paradigm shift where islands are not seen as the ‘end of the colonization road’. They argue that ‘reverse colonization’ is likely for two reasons. First, most island species are direct descendants of successful colonists and should be able to make the trip back, or to somewhere else (although island taxa may loose their dispersal abilities (Darwin, 1909)). Second, even though islands harbor relatively few species, and thus potential dispersers, continents are huge targets.
There are two hypothesized episodes when overland dispersal between the proto-Antillean landmasses and the Americas could occur. The proto-Greater Antilles plate may have connected with North or South America as it passed between them over 60 Mya. A more recent (ca 35 Mya) land connection has also been proposed (the GAARlandia hypothesis (Iturralde-Vinent & MacPhee, 1999)). Moreover, Eastern Jamaica and Cuba are thought to have been linked through proto-Greater Antilles connections to N. and C. America before fragmenting.
The role of dispersal. Layered upon the complex geological history are the inherent properties of the species themselves, such as their dispersal ability. Poor dispersers are less likely to reach isolated islands and thus should be less widespread than better dispersers. However, occurring on more islands does not mean good dispersers must be more diverse across archipelagos. A key question surrounding dispersal is to ‘determine the impact of dispersal distances and ballooning propensity on gene flow and speciation…” (Bell et al., 2005). Gene flow is more rapidly disrupted in poor dispersers because even narrow barriers can be effective isolators. Hence processes of allopatric speciation will start operating earlier in poor dispersers. For example, the spider genus Pholcus, not known to disperse by ballooning, has 25 Macaronesian archipelago endemics. This diversity is a result of a single chance dispersal to the archipelago, followed by diversification within and across its islands (Dimitrov et al., 2008). Similarly, the intermediate disperser Loxosceles is found on very few islands, but has six Caribbean endemics (Table 1). In contrast, Argyrodes spiders are excellent dispersers and have been documented on islands across the Caribbean, yet no Caribbean endemics are currently known (Table 1). Multiple processes may be at work in the case of intermediate dispersers, e.g. Hawaiian thomisid spiders are both diverse and widespread, suggesting that “…dispersal has been sufficiently limited to allow extensive ecological diversification, yet frequent enough that interisland migration is more common than speciation.” (Garb & Gillespie, 2009). The spider genus Modisimus is such an example, occurring on many islands and being highly diverse, e.g. on Hispaniola.
Collecting sites to date:

Puerto Rico (2011 and ongoing)
General: Cambalache State Forest, Guanica State Forest, El Yunque National Forest, Mata de Platano field station, Toro Negro field station, Piñales
Caves: Cueva Clara and Cueva Oscura, Aguas Buenas; Cueva Balcones and Cueva Vientos, Florida; Cueva Matos and Cueva María Soto, Arecibo; Cueva el Convento and Cueva Mapancha, Peñuelas; Cueva Gemelos, Cueva Cangrejos, and Cueva Cabachuelas, Morovis; Cueva 'Corozal', Corozal; Cuevas "Mata de Platano" 1 and 2, Arecibo; Cueva Convento and Cueva Vientos, Guayanilla; Cueva Adrobel, Camuy.
Mona Island (Puerto Rico, 2012 and ongoing)
General: Punta Capitan, Sardinera, Carabinero, Punta Caigo no Caigo, El Faroe, Bajura los Cerezos
Caves: Cueva Doña Geña baja, Cueva Doña Geña arriba, Cueva las Losetas, Cueva Lirio, Cueva Murcielagos, Cueva Caballos, Cueva Caigo no Caigo, Cueva Pozo del Portugues.
Cuba (2012 and ongoing)
General: Turquino National Park, BIOECO field station Siboney, Humbolt National Park, Sierra de Cubitas National Park, Valle de Viñales National Park.
Caves: Cueva de Atabex, Siboney; Cueva de Maguana, El Yunque; Cueva de Lechuza, Cubitas; Cueva de María Teresa; Cubitas; Cueva de Tunel, Viñales
Dominican Republic (2012 and ongoing)
General: Parque del Este National Park, Los Haitises National Park, Isabel de Torres Scientific Reserve, Puerto Plata area schools, Monte Cristi National Park, Valle Nuevo Scientific Reserve, José Armando Bermúdes National Park, Jaragua National Park, Bahoruco National Park, Enriquiollo National Park.
Caves: Cueva del Puente, Cueva Berna, Cueva Sabo, Cueva la Arena, Cueva de Linea, Cueva Guayuyus.
Jamaica: (2013)
General: Hellshire Hills, Blue Mountains National Park, Jackson Bay, Millbank, Cockpit Country, Windsor Research Center.
Caves: Windsor Cave, Hope Gate Cave-Coopers Pan, Daniels Town Cave, Dramalie Cave, Carambie Cave, Harvey Cave 1 and 2, Swanzee Cave, Jackson Bay Caves
USA: (2013, and ongoing)
General: transect from Washington DC down the mountain range to the Florida Keys.
Costa Rica: (2013, and ongoing)
General: La Selva, Barra Honda,
Honduras: Summer 2012
Cusuco National Park (spearheaded by Stuart Longhorn)
Lesser Antilles: (Winter 2013 and ongoing)
Nevis and St Kitts:
Sint Maarten/St Martin: Ile Tintamarre
Saba
Sintt Eustatius
Saint Barthelemy
Martinique
Montserrat
Guadaloupe: Grand Terre, Marie Galante
Dominica
Saint Vincent and the Grenadines: Bequia, Union Island
Saint Lucia
Grenada: Carriacou
Mexico: Summer 2014
Colombia: Summer 2015
WESTERN INDIAN OCEAN
Our work in the Indian Ocean is supported, in part, by National Geographic grants (Global Exploration Fund (GEFNE29-11), Committee for Research and Exploration (8655-09).
The island chains in the western Indian Ocean (Madagascar, Comoros, Seychelles, Mascarenes) have also been identified as Conservation International hotspot of biodiversity. These islands vary vastly in size, age, isolation, and geological formation. The origin of their terrestrial biota is relatively poorly understood, especially that of the Mascarenes (Mauritius, Reunion, and Rodrigues). Madagascar represents a mixture of Gondwanan vicariant lineages and more recent colonizers arriving via Cenozoic dispersal, mostly from Africa (Agnarsson and Kuntner 2011). Dispersal must explain the biota of the smaller islands such as the Comoros and the chain of Mascarene islands, but relatively few studies have pinpointed the source of colonizers, which may include mainland Africa, Asia, Australasia, and Madagascar (Agnarsson and Kuntner 2011). Examples of ancient Gondwanan radiations on Madagascar include boid snakes, podocnemid turtles, and iguanid lizards (Noonan and Chippindale, 2006), typhlopid blindsnakes (Vidal et al., 2010), the extinct elephant birds (Cooper et al., 2001), and cichlid and rainbow fishes (Chakrabarty, 2004; Sparks and Smith, 2004; but, see Ali and Aitchison, 2008). However, existing calibrated phylogenies for vertebrates, invertebrates, and plants are only rarely compatible with the vicariant model (Yoder and Nowak, 2006). Thus, the Cenozoic model seems to apply to more lineages of Madagascar fauna and flora (Yoder and Nowak, 2006), in particular to vertebrates (Vences et al., 2001; Raxworthy et al., 2002; Hume, 2007) and especially mammals (Tattersall, 2006; Masters et al., 2006, 2007; Russell et al., 2008).
The island chains in the western Indian Ocean (Madagascar, Comoros, Seychelles, Mascarenes) have also been identified as Conservation International hotspot of biodiversity. These islands vary vastly in size, age, isolation, and geological formation. The origin of their terrestrial biota is relatively poorly understood, especially that of the Mascarenes (Mauritius, Reunion, and Rodrigues). Madagascar represents a mixture of Gondwanan vicariant lineages and more recent colonizers arriving via Cenozoic dispersal, mostly from Africa (Agnarsson and Kuntner 2011). Dispersal must explain the biota of the smaller islands such as the Comoros and the chain of Mascarene islands, but relatively few studies have pinpointed the source of colonizers, which may include mainland Africa, Asia, Australasia, and Madagascar (Agnarsson and Kuntner 2011). Examples of ancient Gondwanan radiations on Madagascar include boid snakes, podocnemid turtles, and iguanid lizards (Noonan and Chippindale, 2006), typhlopid blindsnakes (Vidal et al., 2010), the extinct elephant birds (Cooper et al., 2001), and cichlid and rainbow fishes (Chakrabarty, 2004; Sparks and Smith, 2004; but, see Ali and Aitchison, 2008). However, existing calibrated phylogenies for vertebrates, invertebrates, and plants are only rarely compatible with the vicariant model (Yoder and Nowak, 2006). Thus, the Cenozoic model seems to apply to more lineages of Madagascar fauna and flora (Yoder and Nowak, 2006), in particular to vertebrates (Vences et al., 2001; Raxworthy et al., 2002; Hume, 2007) and especially mammals (Tattersall, 2006; Masters et al., 2006, 2007; Russell et al., 2008).
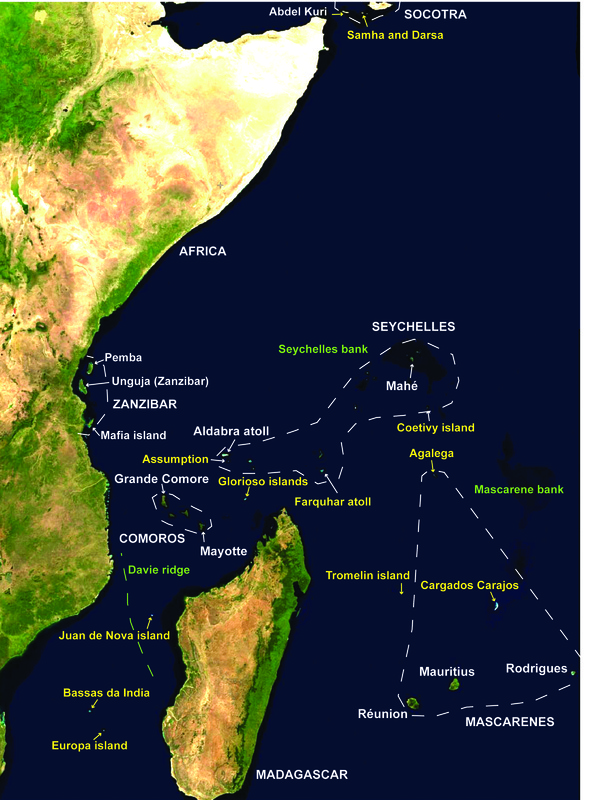
The western Indian Ocean and its many islands
Our island biogeography projects seek to understand reveal the biogeographical history of various spider lineages in the western Indian Ocean, with our focus on the interplay of isolation and dispersal abilities. Our first results (Kuntner and Agnarsson 2011a, in review) from the region, based on Nephilidae are consistent with (1) Cenozoic dispersal to Madagascar from African, and subsequently on to the Mascarenes, and (2) that divergences are shallower among islands in better dispersers, such that intermediate dispersers (Nephilengys, figure below) are most diverse across the archipelagos. However, Nephilidae is a species poor lineage so the generality of these findings remain to be tested with other lineages.
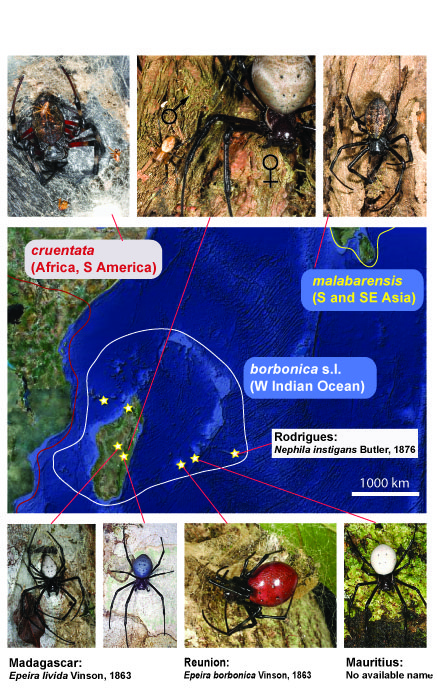
Distribution of Nephilengys species in the region
|

Phylogeny supporting the recognition of three western Indian Ocean Nephilengys species
|
Collecting sites to date
Madagascar, 2001, 2008, 2010: Andasibe National Park, Montagne d'Ambre National Park, Ambohitantely National Park, Masoala National Park, Ranamofana National Park
Comoros Islands, 2008: Mayotte
Mascarene Islands, 2008: Réunion, Mauritius, Rodrigues
Mozambique, 2013: Gorongoza National Park, area around Beira
Seychelles, 2013: Mahe, various localities, Praslin, various localities, Silhouette, various localities, Le Digue, various localities.
Comoros Islands, 2008: Mayotte
Mascarene Islands, 2008: Réunion, Mauritius, Rodrigues
Mozambique, 2013: Gorongoza National Park, area around Beira
Seychelles, 2013: Mahe, various localities, Praslin, various localities, Silhouette, various localities, Le Digue, various localities.
